Overcoming Brain Tissue Complexity: Myelin, Lipids, and Fragile Neuronal Nuclei
Why brain FFPE is different from every other tissue type
Researchers who have processed FFPE tissue from pancreas, liver, or kidney often assume brain tissue will behave similarly. It does not. The brain is roughly 50% lipid by dry weight, wrapped in myelin sheaths that generate massive amounts of non-cellular debris during dissociation. Manual trituration -- the standard approach for breaking apart tissue -- applies force that is simultaneously too gentle to fully clear myelin and too harsh for the large, delicate nuclei of cortical neurons and Purkinje cells.
The practical consequence is a problem that one researcher described as "pick your poison." Gentle processing leaves debris that overwhelms downstream platforms. Aggressive processing clears some debris but breaks the fragile neuronal nuclei you actually want. In either case, robust immune cells -- microglia, macrophages -- survive the prep and dominate the resulting single-nucleus data, while neurons, oligodendrocytes, and astrocytes are under-represented or lost entirely.
This guide walks through each brain-specific challenge, explains why standard FFPE protocols fall short, and describes how automated cartridge-based processing on the Singulator 200+ addresses the biology that makes brain tissue uniquely difficult.
TL;DR - Brain FFPE processing essentials
- Brain tissue generates more myelin and lipid debris than any other FFPE sample type, contaminating nuclei suspensions and wasting sequencing reads
- Manual trituration creates a cell-type bias: robust microglia survive, fragile neuronal nuclei break, and your data skews toward immune populations
- 50-60% of precious postmortem brain tissue is lost during manual processing, with only about one-third of the remainder being intact nuclei
- The Singulator 200+ uses controlled mechanical force and built-in filters to reduce brain-specific debris while preserving fragile cell types
- The two-cartridge FFPE workflow (GREEN then YELLOW NIC+) processes brain tissue from a single curl with less than 5 minutes hands-on time
Brain-specific challenges and how to address them
Five obstacles that make brain FFPE tissue uniquely difficult for single-nucleus workflows, and practical approaches for each.
Myelin debris Understand why myelin generates more debris in brain tissue than in any other organ
Myelin sheaths are lipid-protein structures that insulate axons throughout the central nervous system. In an intact brain, myelin accounts for a large fraction of the tissue mass -- particularly in white matter regions like the corpus callosum, internal capsule, and subcortical fiber tracts. When brain tissue is dissociated, these myelin sheaths fragment into particles that range from sub-micrometer flecks to large aggregates visible under a standard microscope.
The problem is scale. A pancreatic FFPE section might produce debris that constitutes 10-20% of the dissociated material. A brain section -- especially one that includes white matter -- can produce debris that equals or exceeds the volume of intact nuclei. Field evaluation of manual brain FFPE preparations has described the output as "practically an equal mix of nuclei, cell debris, cell ghosts, and micromolecular debris." That ratio is not acceptable for single-nucleus platforms, where debris consumes barcodes, generates ambient RNA background, and reduces the proportion of reads mapping to real cells.
White matter regions (corpus callosum, corona radiata) produce substantially more myelin debris than gray matter regions (cortical layers, hippocampal formation). If your block includes both white and gray matter, expect higher debris loads than blocks cut from pure gray matter. Plan your sectioning accordingly, and communicate with your pathologist about which regions the block contains.
How built-in filtration addresses myelin
The Singulator 200+ cartridges contain integrated filters that separate myelin debris from intact nuclei during processing. Unlike manual protocols, where debris removal requires additional spin-and-wash steps (each of which risks losing nuclei), the on-cartridge filtration happens within the sealed system as part of the standard workflow. The result is a cleaner suspension delivered directly from the instrument, without the repeated centrifugation that causes additional nuclei loss.
For blocks with exceptionally high myelin content, Precision Cell Systems offers a Nuclei Debris Removal Reagent that can be used as a post-processing step after the Singulator 200+ workflow. This provides an additional layer of cleanup for the most challenging white matter-heavy samples, without requiring the 3-5 additional centrifugation steps that manual debris removal protocols demand.
Lipid contamination Address lipid contamination before it compromises downstream platforms
The brain is the most lipid-rich organ in the body. Cholesterol, phospholipids, and sphingolipids are concentrated in neuronal membranes, glial cell processes, and -- most problematically -- the myelin sheaths discussed in the previous panel. When FFPE brain tissue is deparaffinized and dissociated, these lipids are released into the suspension along with the nuclei. Formalin fixation partially crosslinks membrane lipids, but enough is liberated during processing to create visible lipid films on the suspension surface.
Lipid contamination causes two downstream problems. First, lipid droplets and micelles can occlude microfluidic channels on single-cell platforms, leading to reduced cell capture rates or complete channel blockage. Second, lipid-associated molecules contribute to the ambient background signal, muddying the transcriptomic data from captured nuclei. Both problems compound: a preparation with high lipid contamination wastes reagents, produces fewer usable cells, and generates noisier data from the cells it does capture.
Effective deparaffinization removes paraffin wax and a portion of the free lipids simultaneously. The Singulator 200+ GREEN FFPE cartridge uses a proprietary safe solvent for deparaffinization -- no xylene, no CitriSolv, no fume hood. The enclosed cartridge system prevents the solvent evaporation and timing drift that cause incomplete deparaffinization in manual protocols, where leftover paraffin traps lipids and reduces nuclei release.
Why manual deparaffinization fails on brain tissue
Manual deparaffinization protocols typically use xylene or xylene substitutes followed by an ethanol rehydration series. The process works reasonably well for low-lipid tissues. Brain tissue, however, contains so much endogenous lipid that the solvent must contend with both embedded paraffin and tissue-derived lipids simultaneously. Operators frequently report residual waxiness in brain samples that are "fully deparaffinized" by the standard protocol timing. This residual material interferes with enzymatic digestion in the nuclei isolation step, reducing both yield and nuclei quality.
The Singulator 200+ proprietary safe solvent eliminates the need for a fume hood during deparaffinization. This is a practical consideration for labs that share fume hood space or process FFPE samples in facilities without dedicated chemical handling infrastructure. The entire two-cartridge workflow runs at the bench.
Neuronal fragility Preserve fragile neuronal nuclei that manual trituration destroys
Not all nuclei are equally robust. Cortical projection neurons, hippocampal pyramidal cells, and cerebellar Purkinje neurons have large nuclei with open chromatin configurations that make them mechanically fragile. Compare this to microglia, which have compact, dense nuclei designed to survive the physical stresses of immune surveillance. When a pestle or pipette tip applies force to a tissue homogenate, the compact microglial nuclei survive while the larger neuronal nuclei rupture. This is not a subtle effect.
The physics is straightforward. Force applied through a narrow bore pipette tip creates shear stress that scales with particle diameter. Larger nuclei experience proportionally more shear at the same flow rate. Neuronal nuclei can be 10-15 micrometers in diameter -- roughly twice the size of microglial nuclei. At the trituration forces needed to break apart fixed brain tissue, the selective destruction of larger nuclei is essentially guaranteed.
Researchers processing brain FFPE manually face an inescapable trade-off. Gentle trituration preserves some neuronal nuclei but leaves tissue incompletely dissociated, reducing yield and leaving behind clumps that bias the sample. Aggressive trituration improves dissociation but preferentially destroys the large, fragile nuclei of the cell types most relevant to neurodegenerative disease. There is no manual setting that resolves this -- the physics of pipette-based shear stress does not allow selective force application.
Controlled mechanical force as the alternative
The Singulator 200+ applies mechanical force through engineered cartridge geometry rather than operator-dependent pipetting. The force is calibrated, consistent, and distributed across the tissue rather than concentrated through a pipette bore. This controlled processing preserves large neuronal nuclei while still achieving complete tissue dissociation -- a combination that is not achievable through manual trituration regardless of operator skill.
Manual brain FFPE dissociation typically requires 3-5 hours including deparaffinization, rehydration, enzymatic digestion, and trituration -- all in a fume hood. The Singulator 200+ completes the entire workflow in approximately 60 minutes with less than 5 minutes of hands-on time. Beyond the time savings, the shorter exposure to enzymatic and mechanical stress means less cumulative damage to fragile nuclei.
Cell-type bias Recognize and prevent cell-type bias in brain nuclei preparations
Cell-type bias is the downstream consequence of everything described in the previous panels. When myelin debris is not removed, it consumes barcodes that should be assigned to real nuclei. When lipid contamination raises ambient RNA levels, computational decontamination algorithms struggle to assign transcripts correctly. And when neuronal nuclei break during trituration, the surviving population skews toward microglia, macrophages, and other immune cells whose compact nuclei tolerate mechanical stress.
This bias is not just a data quality issue -- it is a scientific problem. If a researcher is studying neuronal gene expression changes in Alzheimer's disease, but the preparation method systematically under-represents neurons, the resulting dataset does not reflect the tissue biology. The researcher is left analyzing the cells that survived the prep, not the cells that matter for the disease.
What cell-type bias looks like in the data
On a UMAP or t-SNE plot from manually processed brain FFPE, microglial clusters tend to be disproportionately large relative to their actual tissue abundance. Neuronal clusters -- particularly excitatory projection neurons -- are small or absent. Oligodendrocyte precursors may be present, but mature myelinating oligodendrocytes (whose nuclei are embedded in myelin sheaths) are often under-represented. The resulting cell-type proportions do not match what spatial transcriptomics or immunohistochemistry shows in adjacent sections from the same block.
If you have access to spatial transcriptomics data (Xenium, MERFISH, Visium) from adjacent sections, compare the cell-type proportions from your single-nucleus preparation against the spatial reference. The Dana Pe'er lab at Memorial Sloan Kettering has used Singulator 200+ nuclei alongside Xenium spatial data for this type of paired validation. Concordance between the two methods indicates that your sample prep is preserving representative cell-type diversity.
Manual FFPE processing loses 50-60% of starting material during the workflow. Of the remaining 40-50%, only about one-third are intact nuclei -- the rest is debris, cell ghosts, and fragments. So from a starting section, roughly 13-17% ends up as usable nuclei, and those nuclei are biased toward robust cell types. The Singulator 200+ shifts this equation by reducing material loss and preserving fragile populations, meaning more of the original tissue biology reaches the sequencing platform.
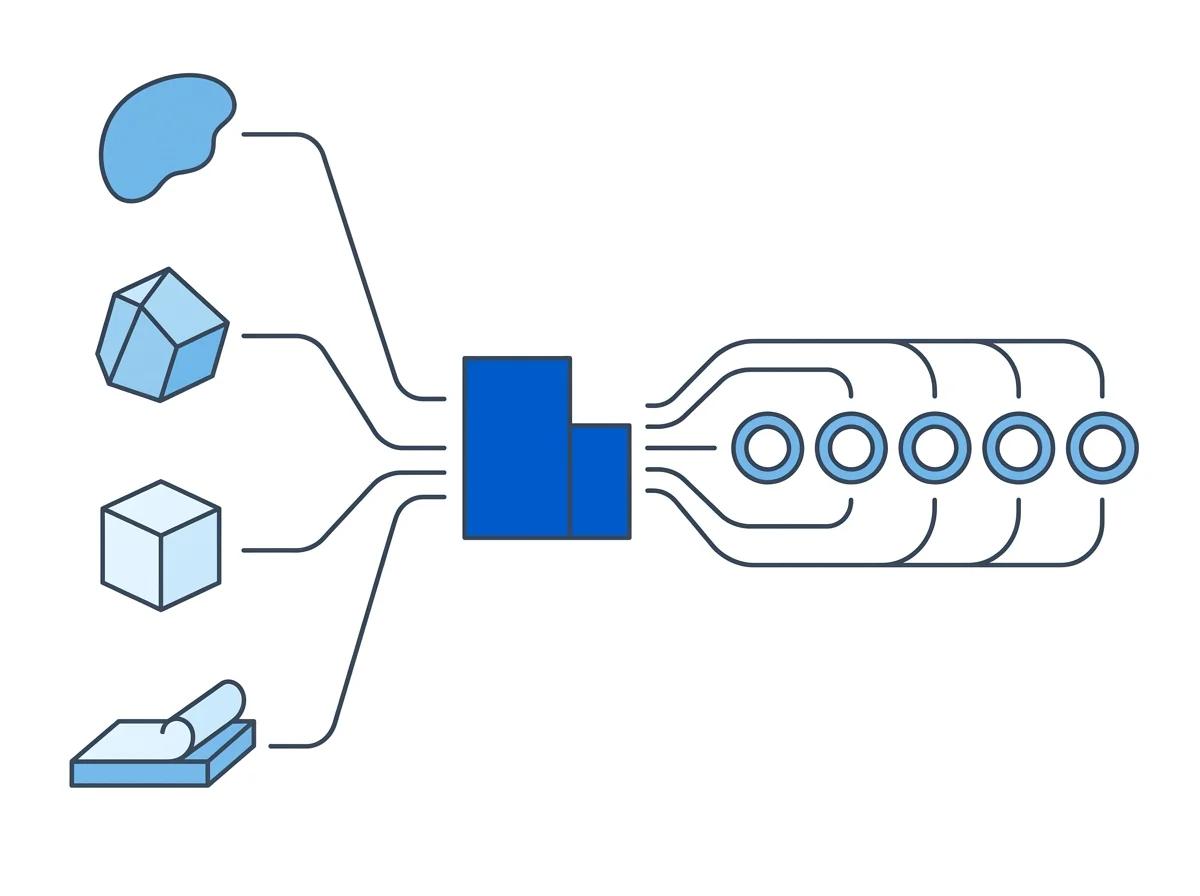
Automated brain processing Process brain FFPE tissue with the two-cartridge automated workflow
The Singulator 200+ FFPE workflow uses two cartridges in sequence to handle the complete process from paraffin-embedded section to sequencing-ready nuclei. Each cartridge is pre-loaded with the reagents and filters needed for its step. The operator loads the tissue, places the cartridge on the instrument, and starts the protocol. The rest is automated.
FFPE two-step workflow S200+ Only
What the automation changes for brain tissue
Three things that manual protocols cannot standardize are standardized by default on the Singulator 200+. First, deparaffinization timing and conditions are identical every run -- no more overnight xylene soaks because someone left for the day. Second, mechanical force is calibrated and consistent, eliminating the operator-dependent trituration that destroys neuronal nuclei. Third, built-in cartridge filters separate myelin and lipid debris from nuclei during processing, rather than requiring post-processing cleanup steps that lose additional material.
The input requirements match the reality of biobank allocations. The Singulator 200+ processes brain FFPE inputs as small as 2 mg of tissue or a single 50 micrometer curl. From a single curl, the instrument consistently yields greater than 1 million nuclei -- far more than the 10,000-20,000 typically loaded onto a 10x Flex chip.
The complete brain FFPE workflow on the Singulator 200+ requires 4 pipetting steps and less than 5 minutes of hands-on time. Compare this to 28 pipetting steps and 25 minutes of hands-on time for semi-automated alternatives, or 3-5 hours total for fully manual protocols. The 81% reduction in hands-on time is not just about convenience -- it is about reducing the number of points where human variability can affect the outcome.
In the PDAC FFPE tissue study, Singulator 200+ replicates yielded 1.0M and 1.0M nuclei -- compared to 1.5M and 0.4M from semi-automated processing of the same tissue. That 3.75-fold improvement in consistency is critical for brain tissue studies, where longitudinal cohorts and multi-site consortia need comparable data across samples processed weeks or months apart.